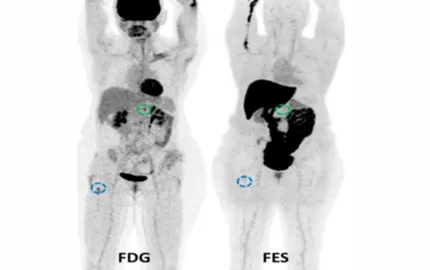
- Accueil >
- Actualités
Les actualités de l'Institut Curie
Découvrez les dernières actualités de l'Institut Curie.
À la une


Rechercher une actualité
Filtrer
Filtrer
Date de publication
Domaines
Rechercher une actualité
567 résultat(s)
- Institutionnel
Rapport annuel 2025 de l’Institut Curie : Inventons un avenir sans cancers incurables
22/07/2026

- Institutionnel
Nouveaux chefs de départements et de services : retour sur les nominations du 1er semestre 2026
23/06/2026